Ist nicht-reduzierbare Komplexität widerlegt?
Eines der bekanntesten Argumente für Schöpfung ist das Argument der nicht-reduzierbaren Komplexität. Hierzu gibt es von atheistischer Seite verschiedene Gegenargumente. Ich möchte hier auf konkrete evolutionstheoretische Entstehungsmodelle für nicht-reduzierbar komplexe Strukturen eingehen. Allgemeine Widerlegungsversuche werden von der Studiengemeinschaft Wort & Wissen zugenüge behandelt.
Der Bombardierkäfer:
Der Bombardierkäfer ist eines der bekanntesten Beispiele für nicht-reduzierbare Komplexität, welches in diesem Artikel ausführlich behandelt wird. Auf talkorigins.org findet sich ein 15-schrittiges Entstehungsmodell, auf welches im Folgenden eingegangen werden soll.
Die angegebenen Schritte sind:
- Chinone werden von Epidermiszellen zur Bräunung der Oberfläche produziert. Es wird darauf verwiesen, dass diese bei vielen Käfern vorkommen.
- Ein Teil der Chinone wird nicht aufgebraucht und bleibt auf der Oberfläche des Käfers zurück, wodurch er ungenießbar wird. Es wird darauf verwiesen, dass Chinone bei einer Vielzahl heutiger Käfer als Verteidigungssekret genutzt wird.
- Kleine Vertiefungen entwickeln sich in der Oberfläche zwischen den Flügeldecken. Durch schütteln kann der Käfer mehr Chinone in die Vertiefungen drücken, wenn sie benötigt werden.
- Die Vertiefungen werden tiefer. Muskeln werden leicht um die Vertiefungen herum bewegt, wodurch es möglich wird die Chinone aus einigen von ihnen auszustoßen. Es wird erwähnt, dass viele Ameisen ähnliche Drüsen am Ende ihres Hinterleibes besitzen.
- Manche Vertiefungen (nun Becken) werden so tief, dass die anderen überflüssig werden und sich langsam zurück entwickeln.
- Bei anderen Insekten treten neben Chinonen auch andere chemische Abwehrstoffe auf. Diese helfen ihnen sich gegen Feinde zu verteidigen, die eine Resistenz gegen Chinone entwickelt haben. Einer dieser Stoffe ist Hydrochinon.
- Die Zellen, die die Hydrochinone absondern, entwickeln sich in mehreren Schichten über einen Teil des Beckens, so dass mehr Hydrochinone produziert werden können. Durch Kanäle zwischen den Zellen können die Hydrochinone aus allen Schichten in das Becken gelangen.
- Die Kanäle werden zu einem Gang, der für den Transport der Chemikalien spezialisiert ist. Die sekretorischen Zellen ziehen sich von der Oberfläche des Beckens zurück und werden schließlich zu einem unabhängigen Organ. Es wird darauf verwiesen, dass dieses Stadium – sekretorische Drüsen, die über Kanäle mit Becken verbunden sind – bei vielen Käfern vorhanden ist.
- Die Muskeln passen sich an und verschließen das Becken, so dass die Chemikalien nicht auslaufen können, wenn sie nicht benötigt werden.
- Wasserstoffperoxid, ein übliches Nebenprodukt des Zellstoffwechsels, wird mit den Hydrochinonen vermischt. Die beiden reagieren langsam, sodass eine Mischung aus Chinonen und Hydrochinonen zur Verteidigung verwendet wird.
- Entlang des Ausgangskanals des Beckens, außerhalb des Ventils, das das Becken nach außen hin verschließt, erscheinen Zellen, die eine geringe Menge an Katalasen und Peroxidasen absondern. Diese sorgen dafür, dass mehr Chinone in den Abwehrsekreten erscheinen. Katalasen kommen in fast allen Zellen vor und Peroxidasen sind auch in Pflanzen, Tieren und Bakterien verbreitet, so dass diese Chemikalien nicht von Grund auf neu entwickelt werden müssen, sondern lediglich an einem Ort gesammelt werden.
- Es werden mehr Katalasen und Peroxidasen produziert, so dass die Entladung wärmer ist und durch den bei der Reaktion entstehenden Sauerstoff schneller ausgestoßen wird. Der Käfer Metrius contractus ist ein Beispiel für einen Bombardierkäfer, der aus seinen Reaktionskammern keinen Strahl, sondern eine schaumige Entladung erzeugt. Durch das Blubbern des Schaums entsteht ein feiner Nebel.
- Die Wände dieses Abschnitts des Ausgangskanals werden fester, so dass sie der durch die Reaktion erzeugten Hitze und dem Druck besser standhalten können.
- Es werden noch mehr Katalasen und Peroxidasen produziert, die Wände verhärten sich und formen sich zu einer Reaktionskammer. Schrittweise werden sie zu dem Mechanismus des heutigen Bombardierkäfers.
- Die Spitze des Hinterleibs wird etwas länger und flexibler, sodass der Käfer seinen Ausstoß in verschiedene Richtungen lenken kann.
Den ein oder anderen mag dieses Modell erstmal beeindrucken, aber bei genauer Betrachtung weißt es erhebliche Schwächen auf. Die folgende Betrachtung der Schritte basiert auf einem E-Mail-Verkehr mit Dr. Boris Schmidtgall:
Zum 1. Schritt: Der Verweis, dass Chinone bei Käfern weit verbreitet sind, sagt noch nichts über den evolutiven Übergang zur heutigen Konstuktion aus.
Zum 2. Schritt: Auch dieses „Stadium“ trägt noch nichts zur Entstehung des heutigen Mechanismus bei.
Zum 3.-6. Schritt: Auch hier wird kein Evolutionsmechanismus erklärt. Der bloße Verweis auf Ameisen mit ähnlichen Drüsen sagt noch nichts über die hypothetische Evolution des Bombardierkäfers aus. Zudem fehlt in Schritt 6 die konkrete Erklärung, wie die neuen Chemikalien entstehen sollen.
Zum 7. Schritt: Die Kanäle werden einfach so als gegeben hingestellt. Es fehlt die Erklärung welche Selektionsdrücke zur Entstehung der Kanäle beigetragen haben sollen.
Zum 8. Schritt: Wieder fehlt die Begründung warum die sekretorischen Zellen von der Oberfläche verschwinden und zu einem eigenen Organ werden sollen.
Zum 9. Schritt: Erst jetzt sollen die Muskeln entstehen, die das Becken dicht verschließen. Demnach müssten die Käfer vorher mit einem ständig auslaufendem Becken gelebt haben, was unsinnig erscheint. Wie sollten sie im Notfall dann überhaupt genug Chemikalien parat haben? Erneut fehlt die Begründung wie und warum sich diese Muskeln entwickelt haben sollen.
Weiter ist bei Bombardierkäfern ein Ventil vorhanden, welches ab einem bestimmten Druck öffnet. Das ist viel mehr als nur ein „Muskel“. Es muss der Druck gemessen werden und eine Rückkopplung zum Öffnen muss vorhanden sein. Diese Details werden nicht mal erwähnt.
Zum 10. Schritt: Wenn Wasserstoffperoxid bei den Zellen ein weit verbreitetes Nebenprodukt ist, ist nicht ersichtlich, warum es erst jetzt auftreten soll. Viel mehr wäre zu erwarten, dass auch die Vorfahren dieses hatten. Zudem ist eine langsame Reaktion von Wasserstoffperoxid und Hydrochinon nutzlos. Sie würden sich langsam neutralisieren und hätten keinen Wert für die Verteidigung.
Nicht ohne Grund bestitzt der Bombardierkäfer heute für Wasserstoffperoxid eine getrennte, spezalisierte Kammer. Es ist ein Zellgift in erhöhter Konzentration. Wie soll es da der Käfer geschafft haben, dass ihm die erhöhte Konzentration nicht schadet?
Zum 11. Schritt: Wieder müsste plausibel gemacht werden, warum die Zellen zur Produktion der Katalasen gerade am Becken entstehen. Dies einfach so zu behaupten ist Spekulation, aber keine Wissenschaft.
Zum 12. Schritt: Es werden mehr Katalasen und Peroxidasen produziert, obwohl das Becken noch nicht belastbar ist. Die Käfer würden sich durch die erhöhten Temperaturen selber töten und somit aussterben.
Zum 13. Schritt: Erst jetzt sollen die Wände fester werden. Demnach hätten die Generationen vorher mit ungeeigneten Wänden leben müssen und wären eventuell unter der Hitze der Reaktion kollabiert. – Wieder bestünde die Gefahr des Aussterbens durch Selbsttötung.
Zum 14.-15. Schritt: Auch wieder bloße Behauptungen ohne Begründungen. Warum sollte sich z.B die Spitze des Hinterteils verlängern und flexibel werden? Zudem wäre interessant, was auf genetischer und chemischer Ebene nötig ist, damit ein fester Stoff flexibel wird.
In Summe behauptet dieses Modell nur. Es fehlt an Begründung und der Benennung konkreter Selektionsdrücke und Entstehungsweisen. Dieses Modell widerlegt die nicht-reduzierbare Komplexität des Bombardierkäfers also nicht.
Der Flagellenmotor der Bakterien:
Der Flagellenmotor einiger Bakterien ist ebenfalls ein sehr bekanntes Beispiel nicht-reduzierbarer Komplexität. In der Fachliteratur wurde bislang interessanter Weise kein konkretes Entstehungsmodell vorgestellt. Nicholas Matzke hat aber auf seiner privaten Website ein konkretes Entstehungsmodell vorgestellt, auf das in der Populärliteratur immer wieder verwiesen wird.
Matzke stellt ein 11-schrittiges Modell auf, mit dem er beansprucht erklären zu können, wie der Flagellenmotor nicht-reduzierbar komplex geworden ist:
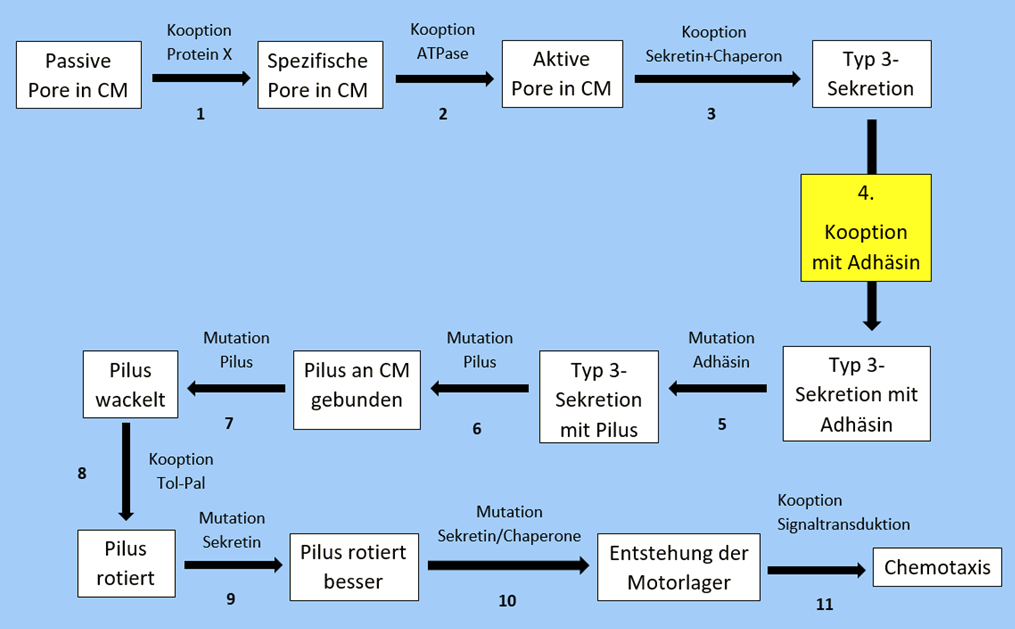
Ich halte mich im Folgenden an die Ausführungen im Buch „Evolution – Ein kritisches Lehrbuch“ (7.Auflage) von Dr. Reinhard Junker und Dr. Siegfried Scherer. Ein paar mehr Informationen zu diesem Modell finden sich in diesem online verfügbaren Artikel von Siegfried Scherer.
Der vierte Schritt – einer der einfachsten – soll hier genauer unter die Lupe genommen werden.
Hier soll eine Kooption (Kopplung / Hinzufügung) eines Adhäsinproteins (Adhäsin befähigt dazu, sich an etwas anheften zu können) an eine Proteinsekretionsmaschine stattgefunden haben. Dafür sind notwendig:
- Präadaptionen (Voranpassungen) des Vorläuferproteins
- eine Duplikation des codierenden Gens
- Umbau zum Adhäsin
- Entstehung einer Kopplungsstelle am Sekretionsapparat
- passende Veränderungen des Sekretionsapparates
- Regulation der Expression (Ablesung und Nutzung der Information in einem Gen) der beteiligten Gene
Es spricht vieles dafür, dass zehn Mutationen nicht reichen, um so eine funktionale Kooption erstmalig hervorzubringen. Der Selektionsvorteil besteht nach Matzke in der Fähigkeit, sich an Öberflächen anheften zu können. Damit diese Struktur Ausgangspunkt für weitere Entwicklungschritte sein kann, müsste sie sich in der Polpulation durchsetzen, wozu ein signifikanter Selektionsvorteil von nöten ist.
Ein wichtiges Problem dieses Evolutionsschrittes ist eben die Kooption eines Adhäsinproteins an einen Sekretionsapparat: Denn dieser Schritt ist nicht ohne Weiteres Ausgangspunkt für die weitere Entwicklung in Richtung eines rotierenden Flagellums. Ist eine neue initiale Funktion mit noch geringem Selektionsvorteil entstanden, führt der Darwin´sche Mechanismus notwendigerweise zur Optimierung dieser Funktion.
Folglich würden die Bindungsstärke der Adhäsionsdomäne an ihr Substrat, eine möglichst feste Verankerung des Adhäsins am Sekretionsapparat und die Proteinsekretion optimiert werden. Diese Optimierung wäre offensichtlich kontraproduktiv für eine Entwicklung zu einem rotierenden Flagellum. Das Adhäsin soll laut Matzke in ein Protein umgebaut werden, das später als Fagellin ein rotationsfähiges Filament mit einem Kanal bildet. Adhäsion hat damit aber wenig zu tun und dementsprechend würde die Optimierung dieser eine Evolution in Richtung Rotationsmotor verhindern.
Der vierte Zwischenschritt in Matzkes Modell eigenet sich also nicht als Zwischenschritt. Wenn wir uns nun klar machen, dass dies noch einer der einfachsten Schritte war, sollte klar sein, dass auch die anderen komplexeren Schritte des Modells nicht viel wahrscheinlicher sind.
Wie bereits erwähnt wird Matzkes Modell in der Fachliteratur nicht diskutiert und auch kein alternatives konkretes Entstehungsmodell vorgestellt. Offenbar sehen auch die Evolutionsbiologen kein plausibles Entstehungsmodell des Flagellenmotors in Sicht.
Somit ist der Flagellenmotor nach wie vor nicht evolutionstheoretisch erklärbar.
Der menschliche Fuß:
Die Längswölbung unseres Fußes besteht aus keilförmigen Knochen. Damit diese Wölbung hält und funktioniert, müssen alle Knochen vorhanden sein. Ausführlicher wird dies in diesem Artikel behandelt.
Manche Atheisten verweisen nun gerne auf Senk- und Plattfüße. Mit denen könne man ja schließlich auch laufen. Zunächst erklären Fehlbildungen nicht die evolutive Entstehung des Fußgewölbes. Sie sind für die nicht-reduzierbare Komplexität also irrelevant. Weiter führen Senk-und Plattfüße zu Beschwerden und Schmerzen, da das Gewicht nicht mehr über die Wölbung verteilt wird. Diese Fehlbildungen bestätigen also im Gegenteil, dass unsere Längswölbung eine Funktion erfüllt.
Zuletzt ist die implizierte Annahme bei diesem Einwand, die Fußwölbung sei überflüssig, ein Schuss ins eigene Knie: Nehmen wir an ein Plattfuß hätte einem Fußgewölbe gegenüber keinen Nachteil. Dann wäre unser Fuß zusätzlich spielerisch Komplex. Unnötige Komplexität ist aus Sicht der ET aber nicht zu erwarten und unplausibel, da die Selektionsdrücke für überflüssige Komplexität fehlen. Sie würde aber zu einem Schöpfer passen, der schlicht Freude an seinem Werk hat.